Актуализация роли спектрального состава света в регуляции биохимического гомеостаза растительной клетки
В рамках совместной работы было проведено комплексное исследовано влияния монохроматического освещения на ростовые и биосинтетические показатели клеток растений, реализующих различные биосинтетические пути, артишока обыкновенного (C. scolymus) и воробейника краснокорневого (L. erythrorhizon). Мы показали, что синий монохроматический свет активирует биосинтез фенилпропаноидов и блокирует синтез изопреноидов, тогда как красный имеет противоположный эффект. Более того, увеличение интенсивности синего светового воздействия приводит к гибели клеток, в то время как красный свет оказывает положительный эффект на рост вне зависимости от интенсивности. На основе полученных данных нам впервые удалось схематизировать роль спектрального состава света в регуляции биохимического гомеостаза растительной клетки, что открывает новые перспективы в светозависимой регуляции распределения клеточных ресурсов.
Рисунок 1. Фитобокс и характеристики освещения. Внешний вид лампы, состоящей из 24 светодиодов (a); ростовые шкафы (б–д) с различными вариантами искусственного освещения (слева направо): тёплый белый и монохроматические источники, такие как красный и зелёный, а также бихроматические источники, сочетающие красный и синий. Представлен нормированный уровень облученности излучения тёплого белого и монохроматических источников света в зависимости от длины волны (нм) (е).
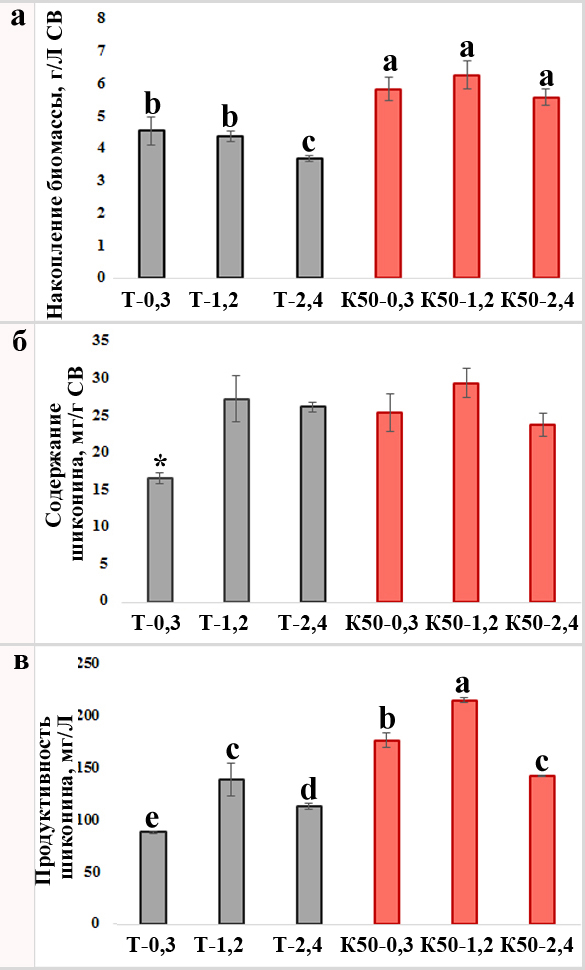
Рисунок 4. Совместное действие красного света и глицерата меди на рост и содержание шиконина в каллусах L. erythrorhizon. Накопление биомассы (a, г/л СВ), содержание шиконина (б) и продуктивность (в) были рассчитаны для каллусов L. erythrorhizon, выращенных в контрольных и экспериментальных условиях. Спектрофотометрический анализ производных шиконина в каллусах L. erythrorhizon, культивируемых в течение 30 дней в условиях Т, темноты и монохроматического красного света с интенсивностью 50 мкмоль×м-2× с-1. Каллусы L. erythrorhizon выращивали на питательных средах с добавлением глицерата меди в стандартной концентрации (0,3 мг/л) и с увеличением концентрации до 1,2 и 2,4 мг/л. Среднее значение ± стандартная ошибка среднего используются для отображения данных из трех отдельных исследований с десятью биологическими повторами. Звездочки или различные буквы над линиями погрешностей указывают на статистически значимые различия (ANOVA, p ˂0,05).
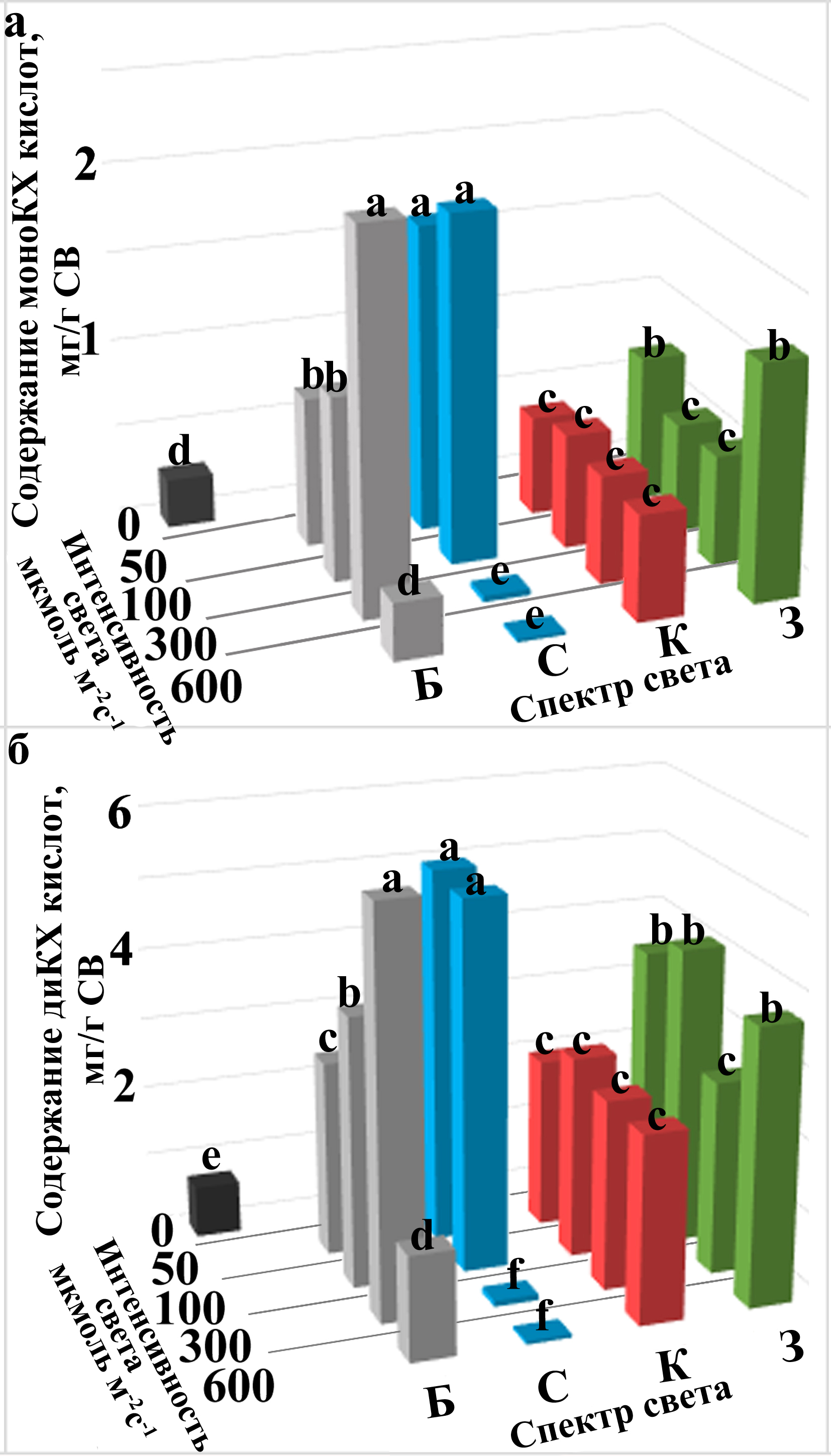
Рисунок З. Содержание кофеилхинных кислот в каллусах С. cardunculus, выращенных при различных условиях освещения. Содержание моноКХ (а) и диКХ кислот (б) в C. cardunculus (мг/г СВ), выращенных в течение 30 дней под различными световыми воздействиями: Т, темнота; тепло-белые, синие, красные и зеленые световые воздействия обозначены как Б, С, К и З, соответственно. Перечисленные световые варианты были использованы с интенсивностью 50, 100, 300 и 600 мкмоль× м-2с-1. Данные, полученные из трех независимых экспериментов с десятью биологическими репликами представлены как средние, а различные буквы над решетками указывают на статистически значимые различия (ANOVA, p ≤ 0,05).
Рисунок 2. Рост культур С. cardunculus, культивируемых при различных световых обработках. Морфология (а) и накопление биомассы (б, г/Л) 30-дневных каллусов C. cardunculus (инокулят 2 г СВ на 50 мл твердой среды), выращенных в течение 30 дней под различными световыми обработками: темнота, теплый белый, синий, красный и зеленый свет обозначены как Т, Б, С, К и З, соответственно. Перечисленные световые варианты были использованы с интенсивностью 50, 100, 300 и 600 мкмоль× м-2с-1. Данные, полученные из трех независимых экспериментов с десятью биологическими репликами представлены как средние, а различные буквы над столбцами указывают на статистически значимые различия (ANOVA, p≤0,05).
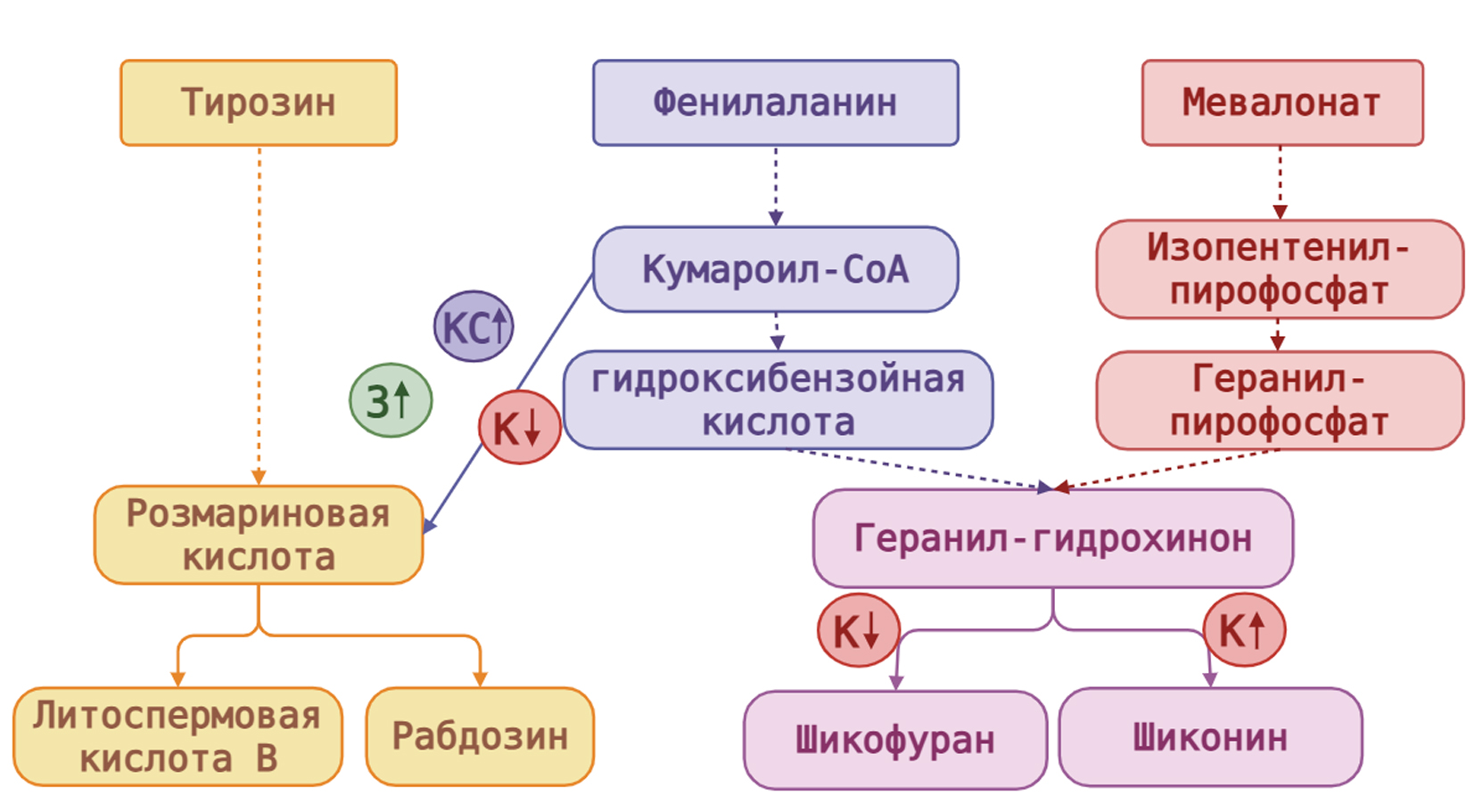
Рисунок 5. Влияние монохроматического и бихроматического светодиодного освещения на вторичный метаболизм в каллусах L. erythrorhizon. Шиконин и шиконофураны являются побочными продуктами геранилгидрохинона. Два ключевых предшественника геранилгидрохинона, геранилпирофосфат (ГПФ), образуются по мевалонатному пути, а п-гидроксибензойная кислота (ГБК) – по фенилпропаноидному пути. Биосинтез ароматического промежуточного продукта ГБК происходит из кумароил-CоА, который является ключевым предшественником для биосинтеза розмариновой кислоты (РК), образующейся из тирозина. Основными производными РК у L. erythrorhizon являются литоспермовая кислота B (ЛКВ) и рабдозин (Sun et al., 2022). Как мы показали в настоящей работе, воздействие монохроматическим зелёным и бихроматическим красным и синим светодиодным светом сдвигало биосинтез со стадии кумароил-СоА на стадию РК, тогда как воздействие низкоинтенсивным красным светом, напротив, сдвигало биосинтез на стадию шиконина. Более того, воздействие низкоинтенсивным красным светом сдвигало биосинтез в сторону шиконина на стадии геранилгидрохинона.